Molecular characterization of Asian hornet paper envelope nests
By Joeri Kaal and Luis Rodríguez-Lado
Abstract
We explored the potential of pyrolysis-gas chromatography-mass spectrometry (Py-GC-MS) as a molecular characterization tool for paper envelop materials in nests of the Asian hornet (Vespa velutina Lepeletier, 1836), a runaway invasive species in Western Europe. The area of study is the NW of the Iberian Peninsula (Galicia, A Coruña province) and the main tree types of the vegetation in the sampled areas are pine, eucalypt and oak, with a smaller abundance of chestnut and fruit trees. The objective was to determine whether molecular fingerprinting in combination with the information on vegetation patterns allows to determine the origin of the decaying wood fibres used for nest construction, and thereby determine the insects’ preferences for raw material. The results confirm that the envelopes consist predominantly of polysaccharides and lignin, and that especially the lignin composition is useful for tracking vegetation sources of raw materials. The relationships between dominant vegetation and nest composition was smaller than expected and there seems to be no clear preference for a given tree type, in spite of very large inter-nest variation in the balance between guaiacyl and syringol lignin products (S/G ranging between 0.0 and 3.0). Thus, the Asian hornet does not exhibit a specific preference for a particular type of vegetation. Instead, it adapts to the available materials in its environment and uses them conveniently in nest construction. This highlights the high flexibility of the Asian wasp in utilizing the resources of the environment it colonizes. However, remarkable intra-nest variation in both colour and S/G ratio indicates that nest colour banding is related to raw material selection. This study contributes to a better understanding of the behavior of V. velutina, which, in turn, helps define appropriate actions to limit its expansion.
Keywords: Asian hornet; paper nest envelopes; molecular characterization; Py-GC-MS; insect ecology.
Towards understanding ecological disaster in the Harz Mountains (Central Germany) by carbon tracing: pyrolysis-GC-MS of biological tissues and their water-extractable organic matter (WEOM)
By Joeri Kaal, César Plaza, Marta Pérez-Rodríguez and Harald Biester
Abstract
Streams and reservoirs in the Harz National Park experience high dissolved organic matter (DOM) concentrations, the cause of which is unknown. We studied potential sources of DOM by means of pyrolysis-GC-MS (Py-GC-MS). The biological materials include vegetation samples (spruce, birch, blueberry, heather, sedge, grass, peat moss, epiphytic moss), microbial sources (epilithic biofilm, lichen, fungi) and excremental fabric. In addition to ground biological samples (bulk organic matter; BOM), their leachates (water extraction of BOM samples followed by filtration; WEOM) were analyzed, to obtain knowledge on solid-liquid transfer effects. Results of BOM showed potential for molecular provenancing on the basis of relative proportions of major biopolymers (lignin, polysaccharides, tannin, resin, etc.) and specific products (biomarkers). Even though WEOM had lower molecular diversity than BOM, identified molecular features of WEOM can be useful to identify potential sources of environmental DOM. Unsubstituted guaiacol prevails among the pyrolysis products of the WEOM of spruce samples (especially wood), which probably originates from pyrolytically decarboxylated vanillic acid moieties. The other plant materials produced a more diverse fingerprint of phenols, guaiacols and syringols, which can be used to distinguish gymnosperm-derived DOM, peatland vegetation and angiosperm trees and shrubs. Several other O-containing moieties such as benzoic acids, benzaldehydes and indanones (not present in BOM) were also frequent products of WEOM. These results lay the foundation for future interpretations of environmental DOM samples’ molecular fingerprints.
Fire, meat and totarol: organic matter in the embankments of the Neolithic site Bastuloken (North Sweden)
By Joeri Kaal, Johan Linderholm and Antonio Martínez Cortizas
Abstract
At the Neolithic site Bastuloken, several subterranean embankments have been identified. The corresponding sediments contain large amounts of bones and lithic elements, indicative of not only massive hunting but possibly also a large (elk) skin processing plant. The present study uses analytical pyrolysis and infrared spectroscopy to track changes in organic matter (OM) composition and relate them to past human activities. It appeared that the sediments contained two layers with a very strong and typical collagen fingerprint (dominated by pyrrole and diketodipyrrole), confirming ubiquitous presence of bone and/or meat. Also, black carbon was abundant in several layers, which is probably an indication of fireplaces. Thus far, the results fit with the hypothesis of major inputs of animal tissue especially during two phases recorded between 30-45 cm and 60-65 cm depth, the first of which also contains higher inputs of totarol, indicative of the presence (and probably use) of resin from Cupressaceae (gymnosperm tree). The level of OM preservation in these Neolithic structures is extraordinarily good and shows huge potential for molecular fingerprinting and deepen our understanding of the activities at this important site and the temporary and spatial variability therein.
Analytical pyrolysis in marine environments revisited
By Joeri Kaal
Abstract
This contribution outlines applications of analytical pyrolysis techniques (APTs) –in particular pyrolysis-gas chromatography-mass spectroscopy (Py-GC-MS) and thermally assisted hydrolysis and methylation (THM-GC-MS)– to marine geochemical research. It is subdivided into two main lines, i.e. the results obtained from (1) sedimentary, particulate and dissolved OM from open water environments (estuaries, seas, oceans), and (2) coastal vegetated habitats such as saltmarsh, mangrove and seagrass ecosystems (Blue Carbon). The main aim is to re-familiarize the marine research community with the possibilities of these infra-utilized techniques for the identification of sources and assessment of degradation/preservation state of macromolecular organic matter.
Featured image: sea surface of the South Atlantic Ocean (near Namibia), from the FUNAMOX 2019 expedition (courtesy of Dr. Peter Kraal, NIOZ).
Naturally halogenated organic matter in Atlantic rankers is concentrated in microbial rather than pyrogenic moieties
By Joeri Kaal and Antonio Martínez Cortizas (Earth System Science, GI-1553, Universidade de Santiago de Compostela, Santiago de Compostela, Spain)
Abstract
Organohalogens in the environment are a booming research subject, due to their role in many ecosystem functions and ecotoxicology. A recent study showed that a series of colluvial soils in NW Spain, known as Atlantic rankers and having the idiosyncratic feature of high loads of aged pyrogenic soil organic matter (OM) from anthropogenic palaeofires, contain large amounts of organically-bound natural bromine. Furthermore, it appeared that the Br/C ratio, indicative of biobromination status of the OM, increases steadily with age during the Holocene. We compared the organobromine record with OM composition as determined by pyrolysis-GC-MS. It appeared that negative excursions in the Br/C record coincide with peaks in pyrogenic matter content and positive excursions with peaks in the proportion of pyrolysis products from microbial constituents. Hence, in addition to age, SOM composition has a secondary controlling effect on organobromine concentrations. And the fact that the Br is associated with microbial SOM is strong evidence of a biogenic source (microbial halogenation), as abiotic surface binding would cause enrichment of Br in the pyrogenic SOM fraction, which has the largest specific surface area.
Molecular properties of soil organic matter in dark buried colluvium from South Germany show abundance of fire residues from Early Neolithic vegetation clearance and slash and burn agriculture
By Joeri Kaal and Sonja Mailänder (Institute of Geography, University of Stuttgart)
Title image in slider: Berndt Lindholm (1841–1914), Slash-and-burn in Idensalmi (Svedjeland i Idensalmi, Savolaks), from Zacharias Topelius (En resa i Finland, Helsingfors Bokförlaget Atlantis, Stockholm 2013) (https://commons.wikimedia.org/wiki/File:B_Lindholm_Svedjeland_i_Idensalmi.jpg)
Geoarchaeological surveys in South Germany (Nördlinger Ries area) showed continuous presence of human occupations since the Palaeolithic and remarkable colluvial soils containing dark buried layers dated to the Neolithic. Here we discuss molecular properties of soil organic matter (SOM) from three of these colluvia by Py-GC-MS. The pyrolysis products could be ascribed to four different SOM components: labile SOM (fresh lignin and polysaccharides), a recalcitrant aliphatic fraction, degraded/secondary SOM (including chitin and degraded carbohydrates) and pyrogenic SOM (Black C, from palaeofires). The Black C was recognised unambiguously from the full range of diagnostic compounds, i.e. abundant polycyclic aromatic hydrocarbons (PAHs) such as naphthalenes, biphenyl, fluorene, phenanthrene, anthracene and pyrene, and N-containing compounds such as benzonitrile, (iso)quinoline and naphthalene carbonitriles. Black soil layers generally contained larger proportions of pyrogenic material but SOM dynamics of these soils seem controlled by intense degradation, eliminating most components of fresh plant material and producing newly-formed degraded SOM (including chitin from fungi and/or arthropods) and a relative accumulation of aliphatic SOM. Nonetheless, the black colour of the Neolithic soil layers is clearly caused by the strong pigmentation of fire residues and reflects a series of (forest) fire events ca. 6000–7000 cal BP. This study shows the potential of Py-GC-MS to identify pyrogenic SOM and simultaneously identify the processes that control SOM dynamics in prehistorical anthropogenic colluvium.
1. Introduction
Pyrolysis-GC-MS is increasingly used for Black C identification and characterization in archaeological sites but also off-site anthropogenic dark earths such as worldwide Terra Preta deposits and colluvial deposits, despite the potential difficulties in distinguishing artificial thermal modifications (during analytical pyrolysis) and the “natural” thermal impact by fire. While many of the major pyrolysis products of Black C, e.g. benzene, toluene, C2-benzenes, pyridine and PAHs, are not specific of Black C, they tend to be enriched in Black C-rich soil and, if the chromatogram quality is optimized, accompanying compounds such as benzene carbonitriles or naphthalene carbonitriles may allow for a definitive identification of the pyrogenic SOM signal.
Excavations and geomorphological research in the study area by Sonja Mailänder and her colleagues Prof. Dr. Wolf-Dieter Blümel and Dr. Joachim Eberle showed a series of partially black-coloured colluvial deposits that formed the basis of a landscape reconstruction exercise, i.e. the PhD research of Sonja Mailänder. For details of this research, the study area, and the deposits we examined by analytical pyrolysis, please refer to Mailänder et al. (2010, 2011).
Aiming to elucidate soil formation processes and to seek for indications of human activities (anthropogenic remains and land use strategies), we performed Py-GC-MS to the SOM from the three soil profiles, with special interest in the nature of the SOM in the buried dark layers.
2. Methods and materials
2.1 Study area, background, sampling
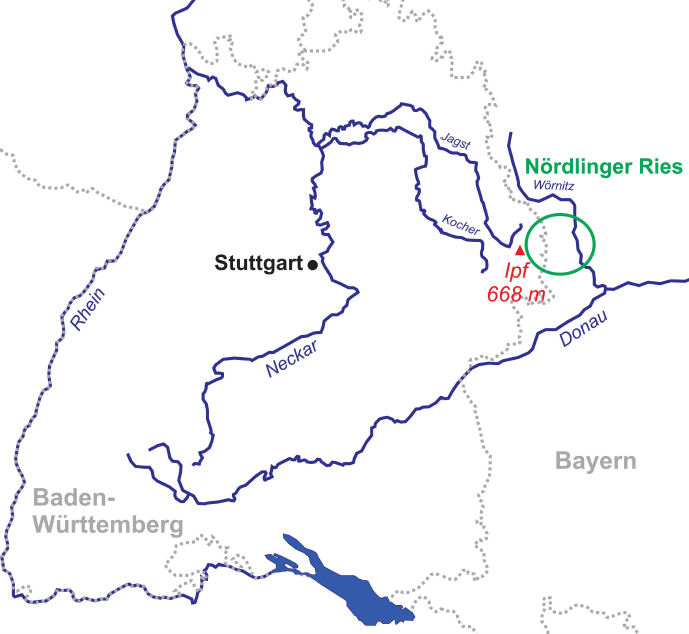
The study area is located nearby the Ipf Mountain (Nördlinger Ries, South Germany, 80 km east of Stuttgart), which is on the border of a crater that formed upon a Miocene meteorite impact 15 million years ago (Figures 1 and 2). Figure 3 shows the colluvial deposits (taken from a pipeline trench), samples analysed, and radiocarbon dates.
The study area is situated in southwest Germany at the western border of the Nördlinger Ries, which is a crater formed by a Miocene meteorite impact 15 Million years ago.
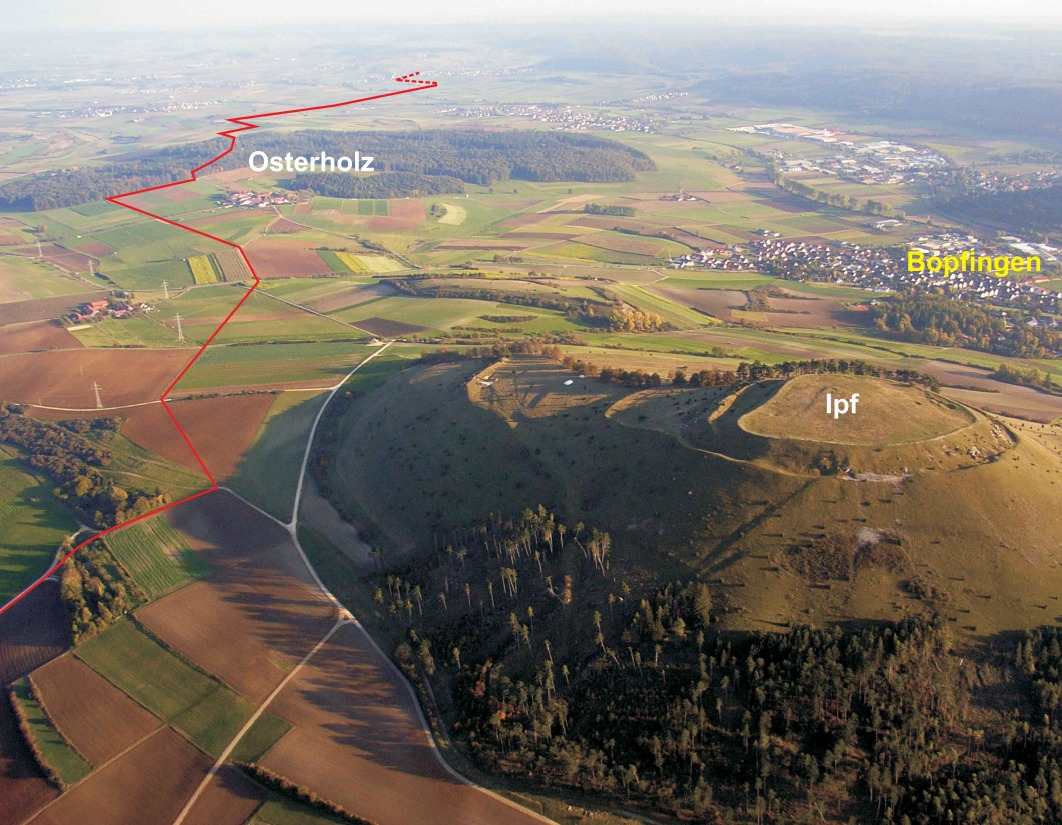
Figure 2. This aerial picture shows the mountain Ipf near the city of Bopfingen. The red line marks the course oft the pipeline ditch, which was investigated in the years 2006 and 2007 (Foto: S. Mailänder, Oktober 2007).
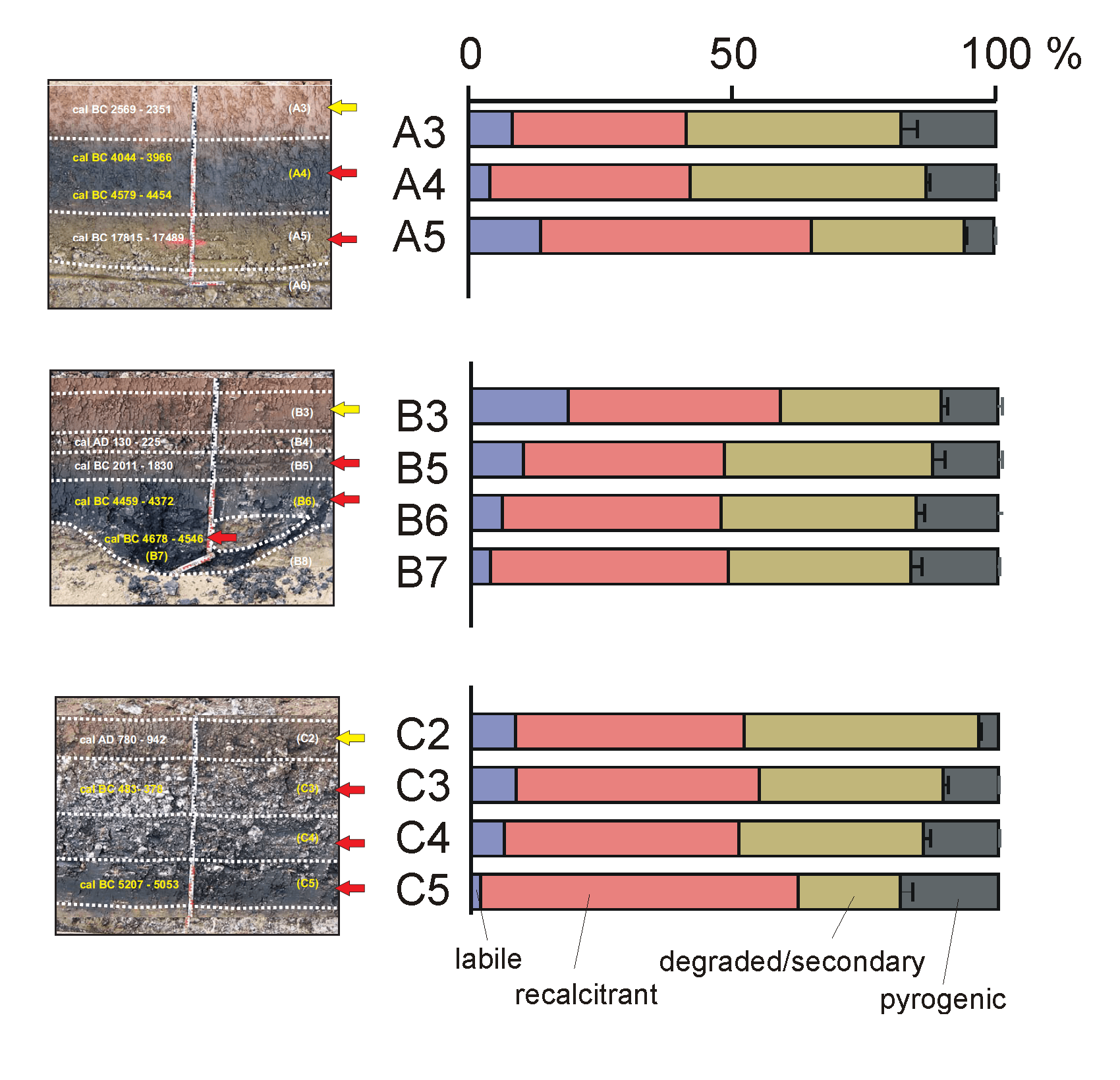
Figure 3. Photographs from the colluvial deposits, taken from a pipeline ditch. Samples from profile A, B and C that were analysed by Py-GC-MS are marked with arrows.
Profile A is located to the north-east of the Ipf hill in the upper reaches of the Goldbach Valley. This valley has no watercourse anymore and is used as grassland and grazed by cattle. In the pipeline trench the black layer (A4), which was covered by colluvial deposits, appeared in the deepest part of the depression and lashed out to the left and to the right. A calcareous-rich layer was found underneath layer A4 (A5). The geological substrate is composed of the Upper Aalenian (Aalenian 2, al2E, former Brown Jurassic β).
Profile B is situated further east within the outer ring of the Ries crater, nearly on top of a small depression, which runs to the Goldbach Valley from the west. Today this area is used for agriculture. The underlying bedrock is a dislocated Upper Jurassic megablock. In profile B a dark layer (B6) was found as well, at the deepest part of the depression lashing out to the slopes and covered by colluvial deposits. In the position of the investigated profile in the pipeline trench, a small pit-like structure appeared underneath (B7).
Profile C was studied on the south-facing, gently tapered slope of the Eger Valley, also within the megablock zone of the crater. Here the basement is built of Bunte Breccia that is a product of the meteorite impact. It consists of multicoloured chaotically mixed rock fragments, which originate from different bedrock formations. On top of a very clayey black horizon (C6), poor in coarse particles, two further dark layers (C5 and C4) are found, which contained many pieces of Upper Jurassic limestone.
2.2 HF treatment
Minerals that catalyze secondary rearrangements during pyrolysis were eliminated by HF treatment. Briefly, 2.0 g of sample were transferred into 50 ml centrifuge tubes and 10 ml of 1 M HCl were added repeatedly to remove carbonates. After the reaction was completed, H2O was added until the 25 ml mark and the resulting suspension was agitated, centrifuged (2500 rpm for 10 min) and the supernatant discarded. Next, 2 % HF solution was added until the 25 ml mark and shaken for 24 hrs, followed by centrifugation and decantation of the supernatant. This HF treatment was repeated 5 times, after which the procedure was executed three times with H2O to wash away the acid and silicates. Finally, the residue was dried at 35 ºC.
2.3 Pyrolysis-GC/MS
Pyrolysis-GC-MS was performed using a Pt filament coil probe resistive heating pyrolyzer (Pyroprobe 5000) from CDS Analytical Inc (Oxford, USA). The pyrolysis interface was connected to a 6890 GC and 5975 MSD from Agilent Technologies (Palo Alto, USA). Approximately 1 mg of sample (HF-treated residue) was embedded in fire-polished quartz tubes using glass wool. Pyrolysis was performed at 750 ºC (750 ºC setpoint temperature is approximately 670 ºC real pyrolysis temperature which is still very high, but found ideal for Black Carbon characterization by Kaal et al. 2009) for 10 s at a heating rate of 10 °C ms-1. The pyrolysis interface and GC inlet were set at 325 °C. The GC oven was heated from 50 to 325 °C at 20 °C/min and held isothermal for 5 min. The GC-MS transfer line was held at 325 °C, the ion source (electron ionisation mode, 70 eV) at 230 °C and the quadrupole detector at 150 °C. The GC was equipped with a (non-polar) HP-5MS 5% phenyl, 95% dimethylpolysiloxane column (ca. 30 m x 0.25 mm i.d.; film thickness 0.25μm). Helium was used as carrier gas (constant flow, 1 ml/min). Two replicates were obtained for each sample. The ca. 50 largest peaks of the total ion current of each pyrolysis chromatogram were listed, giving rise to a compound list of 114 pyrolysis products. The relative proportions of the pyrolysis products were calculated from the peak area of their dominant and/or characteristic mass fragments. Relative proportions are expressed as the percentage of total quantified peak area (% TQPA). Two replicates were obtained for each sample.
3. Results and Discussion
3.1 Py-GC-MS products and source allocation
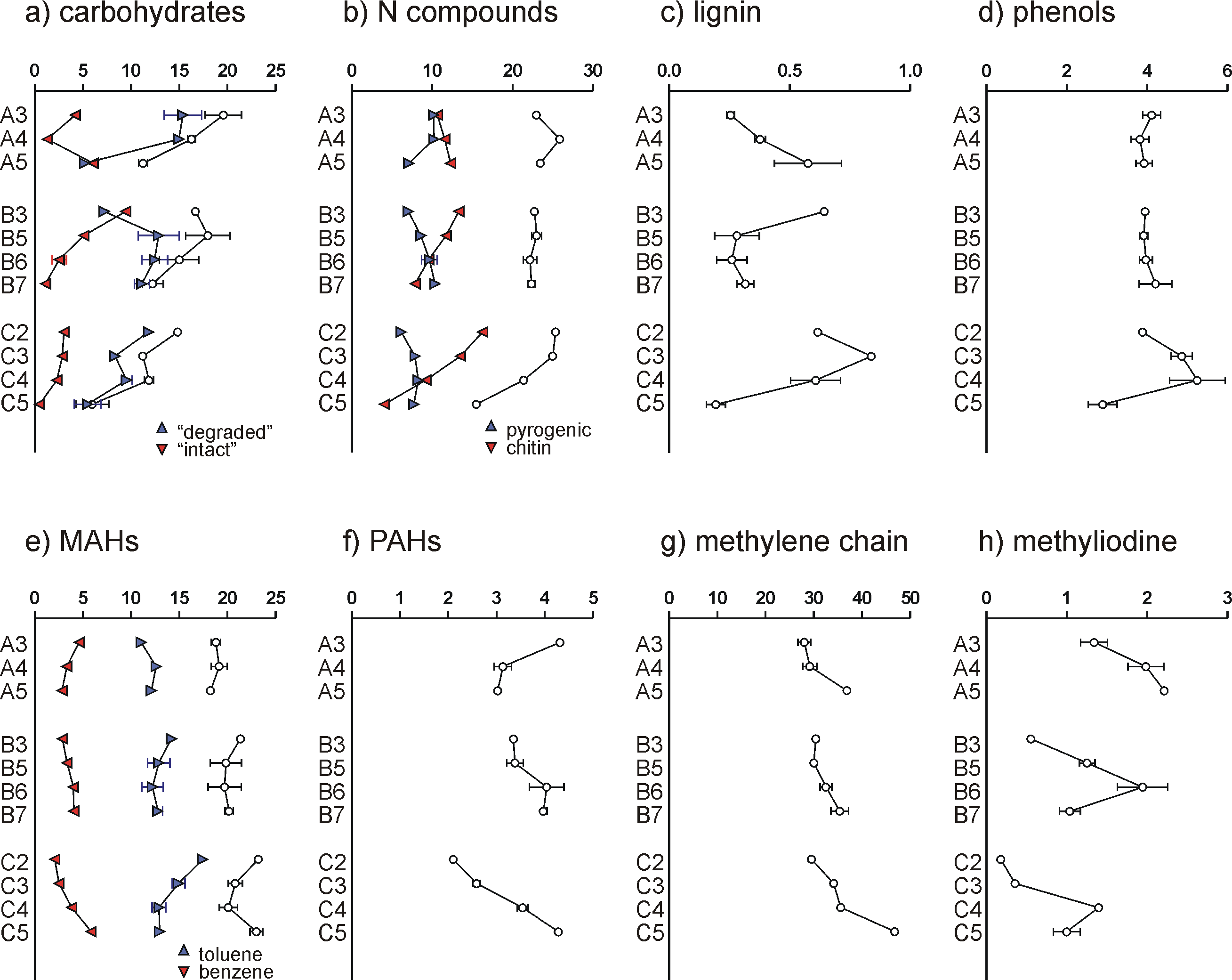
The list of compounds is provided in Table 1 at the bottom of this article. The pyrolysis products were grouped based on chemical structure and/or most likely origin. The markers of carbohydrates are 3/2-furaldehyde, 5-methyl-2-furaldehyde, 4-hydroxy-5,6-dihydro-(2H)-pyran-2-one, dianhydrorhamnose, levoglucosenone and levoglucosan. Of these compounds, levoglucosan, dianhydrorhamnose and 4-hydroxy-5,6-dihydro-(2H)-pyran-2-one originate mainly from relatively intact polysaccharides while levoglucosenone and the furfurals originate from degraded carbohydrates, even though there is no strict division between these two subgroups (intact polysaccharides also produce furfurals upon pyrolysis). An unidentified compound producing base ion m/z 116 was grouped here as it is possibly an indication of moss-derived carbohydrate (Schellekens et al., 2011), and was correlated to the three markers of intact polysaccharide (3x P<0.001). The seven carbohydrate products combined account for 12 ± 4 % of TQPA (Fig. 4a). This proportion decreases with depth in the three soils, which may be explained by preferential carbohydrate decomposition with time. In profiles B and C, the relative proportion of intact carbohydrate markers declines with depth, reflecting carbohydrate degradation. However, in profile A, sample A5, which is the only Late-Pleistocene/Early Holocene sample of the present study, produced a relatively large proportion of intact polysaccharide markers, probably because this layer formed under conditions with low microbial activity (cold and dry).
Figure 4. Relative proportions of the different groups of pyrolysis products. Sum of total compounds in group in white symbols, subgroups are indicated for carbohydrates, N-compounds and MAHs (monocyclic aromatic hydrocarbons).
Pyrolysis products containing one or more atoms of N (N-containing compounds) are abundant, i.e. 23 ± 3 % of TQPA (Fig. 4b). These compounds have a diverse origin, and a VARCLUS cluster analysis using Tanagra software was performed to arrange them into subgroups. Two main clusters were obtained, accounting for 85 ± 4 % of total N-containing pyrolysis products. One cluster consists of (in decreasing order of cluster correlation coefficient) acetamide, C1-pyrrole, a compound with base ion m/z 94, benzeneacetonitrile, trianhydro-2-acetamido-2-deoxyglucose, 2,5-diketopiperazine, C1-pyridine, a compound with base ion m/z 109 (and m/z 66 and m/z 80 as daughter ions of two co-eluting isomers), indole and pyrrole. The compounds m/z 94, m/z 109 and trianhydro-2-acetamido-2-deoxyglucose are markers of the N-acetylglucosamine-based biopolymer chitin, which occurs in invertebrate/arthropod exoskeleta, fungi cell walls and also in molluscs. The diketopiperazine has been identified in pyrolyzates of chitin-entangled protein, and the other products can also be produced by chitin but are less specific. Therefore, the compounds in this cluster are thought to originate primarily from chitin. Chitin seems to be present in all samples (Fig. 4b). Its proportion declines with depth in profiles B and C. An unidentified compound with fragment ions m/z 59, 114 and 204, which could correspond to 1,6-dianhydro-2-acetamido-2-desoxyglucose, was identified in an isolated snail shell fragment from sample A4 (not shown). This compound, which could be a marker of mollusc chitin, was not found in the HF-treated samples. Nonetheless, it is difficult to decide on the source organism of the chitin markers. We believe that they are mainly arthropod-derived and/or fungal, either way being an indication of biological recycling of plant-derived SOM. The second cluster consists of two naphthalene carbonitrile isomers, benzonitrile, N-methylpyrrole, C1-benzonitrile and pyridine. The naphthalene carbonitriles (or naphthonitriles) and benzonitriles are markers of pyrogenic N (Kaal et al., 2009), and pyridine is a less-specific of it (e.g. Kaal and Rumpel, 2009). The depth trend of this pyrogenic N (or Black N; Knicker, 2010) is discussed below. The position of the N-methylpyrrole in this cluster cannot be explained (it is an unlikely product of pyrogenic material). Furthermore, we identified a set of phthalimides (three compounds), which we think are also markers of pyrogenic N. Diketodipyrrole (a dimerization product of protein pyrolysis), C16– and C18-alkylamide, and C16– and C18-alkylnitrile, were also detected but not associated with one of the aforementioned clusters.
In the present study, the guaiacyl- and syringyl-based pyrolysis products of lignin were scarce (0.5 ± 0.2 % of TQPA). Also, only guaiacol and 4-vinylguaiacol were detected. This implies that either lignin was not a significant source of the SOM or that it was profoundly remediated. Lignin is enriched in the SOM of the Early Holocene sample A5 (confirming the preservation of some lignocellulosic source) while it decreases with depth in profiles B and C, probably because of decay (Fig. 4c).
Considering the presence of lignin as well as pyrogenic matter and abundant degraded and secondary biomass, which all produce phenols upon pyrolysis, the phenols (4.1 ± 0.7 % of TQPA) cannot be ascribed to a specific SOM component. From the relative proportion of phenols in profile C (Fig. 4d), which shows a similar trend to that of the lignin markers, it may be concluded that the phenols in that soil originate primarily from degraded lignin. Analogously, in profiles A and B the phenols may be derived mainly from N-containing proteinaceous biomass.
Monocyclic aromatic hydrocarbons (MAHs) account for 24 ± 2 % of TQPA. Toluene is the most abundant product, followed by benzene. These two compounds exhibit inverse depth trends (Fig. 4e). While toluene is correlated to the products of chitin (r2=0.31, P<0.05), benzene is correlated to pyrogenic N (r2=0.48, P<0.001). This behaviour may be expected from soil samples with large proportions of both proteinaceous biomass and pyrogenic matter. The other MAHs (xylenes and ethylbenzene) may have diverse origins, so that they cannot be allocated to a specific SOM component.
Polycyclic aromatic hydrocarbons (PAHs) account for 3.4 ± 0.7 % of TQPA. Here, the main source of these compounds is pyrogenic matter, i.e. Black C. PAHs increase with depth in soils B and C, but not in soil A (Fig. 4f).
The group of methylene chain compounds (MCC) includes homologous series of n-alkanes (C10-C33), n-alkenes (C10-C31), n-fatty acids (C16 and C18) and n-methylketones (C23, C25, C27 and C29), in addition to prist-1-ene. Combined, these compounds account for 34 ± 5 % of TQPA (Fig. 4g). The vast majority of this group can be ascribed to n-alkane/n-alkene pairs (30 ± 5 % of TQPA). The increase with depth in all soils can be explained by the resistance of aliphatic material during SOM degradation, in this case their relative preservation compared to mainly carbohydrates but also compared to chitin.
Other compounds (3.1 ± 1.4 % of TQPA) include methyliodine (m/z 142 and 127; 1.2 ± 0.7 % of TQPA), which is not frequently found in SOM pyroyzates (Fig. 4h). A possible source of this compound is halogenation: these clay-rich soils release significant amounts of I upon weathering which, in combination with the high microbial/biological activity, forms organo-iodine complexes through biotic halogenation. We checked for the presence of methylbromine (which should be formed through the same process; Martínez-Cortizas et al., 2016), and we identified this compound as well (m/z 94, 96). Quantification of Me-Br, not presented in the Appendix because it was not recognised in the initial scannings of the pyrograms, showed that it is correlated to Me-I (r2=0.72, P<0.001), with the relative proportion of Me-I and Me-Br being roughly 4:1. Other compounds detected are an aliphatic pyrolysis product with characteristic fragment ions m/z 83 and 280, frequently associated with fresh plant remains, and an unidentified polar compound with base ion m/z 135 (possibly from chitin).
3.2 Coupling of Py-GC-MS fingerprints and geo-archaeological information
It appeared that the SOM of the soils studied is a mixture of relatively intact plant-derived “labile” SOM (polysaccharides and lignin), aliphatic “recalcitrant” SOM (C10-C25n-alkane/n-alkene pairs), “degraded/secondary” SOM (furfurals, chitin markers) and pyrogenic SOM (benzene, PAHs, benzonitriles, naphthalene carbonitriles), in varying relative proportions. Figure 5 shows the relative proportions of these groups, omitting all compounds of unspecific origin. It seems that the majority of the SOM originates from biotically (microbially or arthropodically) altered SOM (degraded/secondary SOM) and aliphatic “recalcitrant” SOM. This can be explained by the intensity of SOM degradation, eliminating most plant-derived labile components leaving the degraded/secondary (e.g. chitin) and recalcitrant SOM. Preferential decomposition of chitin may explain the relative increase in the proportions of pyrogenic and recalcitrant SOM with depth (profiles B and C). Note that these are only rough estimations that may be severely biased (e.g. because of low pyrolysability of pyrogenic matter).
Figure 5. Relative proportions of labile plant-derived SOM, microbial SOM, recalcitrant aliphatic SOM and pyrogenic SOM.
Profile A. Sample A5 corresponds to the Early Holocene (a post-glacial deposit ca. 14 ka old) and therefore contains no or very few fire residues. The fire regime had not started yet and the SOM is composed predominantly of recalcitrant aliphatic domains (from roots for example). Sample A4 is deep black in colour and has larger microbial and pyrogenic SOM contents. This corroborates the abundance macroscopic charcoal and suggests that the “black particles” mentioned by Mailänder et al. (2010) (referring to sample A4) indeed originate from fire residues, presumably from anthropogenic fire clearance during the Neolithic (upper and lower boundaries dated to 6000 and 6500 cal BP, respectively). Sample A3 (4500 cal BP) is an exception to the rule that the dark-coloured samples have highest Black C contents. It has a reddish colour and is clearly post-Neolithic. This may be related to its low C content (weak pigmenting effect of pyrogenic SOM) and/or the presence of Black C at the lower boundary between A3 and A4.
In profile B, a clear increase in pyrogenic SOM content is observed, culminating into the darkest levels that were dated to 6500 cal BP. Surface levels of profile B have a relatively high proportion of well-preserved plant remains.
A similar pattern is recognised in profile C. The youngest samples analysed here date to 1200 cal BP (sample C2, ca. 800 A.D.) and is a greyish layer that was formed well after the dark-coloured layers that formed more than 1000 years earlier (C3, 2500 cal BP). Sample C2 has no or very little pyrogenic SOM, whereas C3 has slightly higher levels. The highest proportion of pyrolysis products of Black C are found in samples C4 (not dated) and C5 (7000 cal BP), which are also deep black in colour.
4. Conclusions
The geo-archaeological survey already showed clear consistency in the depth-age records and determined age of the dark-coloured deeper layers: they correspond to a rather narrow age range between 6000 and 7000 years ago, corresponding to Neolithic fire events. It has now been established on the basis of molecular data that the black colour of these layers can indeed be safely ascribed to the abundance of Black C, i.e. fire residues. Clearly, considering their thickness, these dark layers correspond to a very intense recurrent fire regime. Early human-induced anthropogenic erosion processes could thus be dated as far back as the early Neolithic. According to Mailänder et al. (2010) it was probably caused by vegetation clearance and initial slash and burn agriculture. The abrupt shift in lighter materials after the Neolithic can be related to the absence of biomass (fuel) after deforestation: post-Neolithic sporadic burning of herbaceous vegetation in pasturelands or elimination of weeds from a fully converted agricultural landscape, of which evidence exists near Ipf, would not give rise to deep-black charcoal-rich colluvium.
REFERENCES
Kaal, J., Nierop, K.G.J., Martínez Cortizas, A., 2009. Characterisation of aged charcoal using a coil probe pyrolysis-GC/MS method optimised for Black Carbon. Journal of Analytical and Applied Pyrolysis 85, 408-416.
Kaal, J., Rumpel, C., 2009. Can pyrolysis-GC/MS be used to estimate the degree of thermal alteration of black carbon? Organic Geochemistry 40, 1179-1187.
Knicker, H., 2010. “Black nitrogen” – an important fraction in determining the recalcitrance of charcoal. Organic Geochemistry 41, 947-950.
Mailänder, S. Eberle, J., Blümel, W.D., 2010. Kolluvien, Auelehme und (An)moore im Umfeld des frühkeltischen Fürstensitzes auf dem Ipf – Ein Beitrag zur Geoarchäologie und Landschaftsgeschichte am Westrand des Nördlinger Rieses. In: Krausse, D.L., 2010, Frühe Zentralisierungs- und Urbanisierungsprozesse. Zur Genese und Entwicklung frühkeltischer Fürstensitze und ihres territorialen Umlandes. Kolloquium des DFG-Schwerpunktprogramms 1171.
Mailänder, S., Hecht, S., Eberle, J., Blümel, W.D., 2011. Geoarchäologische Erkundungen in zwei Muldentälchen östlich des Ipfs am Westrand des Nördlinger Rieses (Süddeutschland). Tagungen des Landesmuseum für Vorgeschichte Halle, Band 6, 129-144.
Martínez Cortizas, A., Ferro Vázquez, C., Kaal, J., Biester, H., Costa Casais, M., Taboada Rodríguez, T., Rodríguez Lado, L., 2016. Bromine accumulation in acidic black colluvial soils. Geochimica et Cosmochimica Acta 174, 143-155.
Schellekens, J., Buurman, P., Fraga, I., Martínez-Cortizas, A., 2011. Holocene vegetation and hydrologic changes inferred from molecular vegetation markers in peat, Penido Vello (Galicia, Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 299, 55-69.
Sources of organic detritus under mussel farms in the Ría de Ares-Betanzos
By Joeri Kaal and CSIC-PROINSA Mussel Lab Team
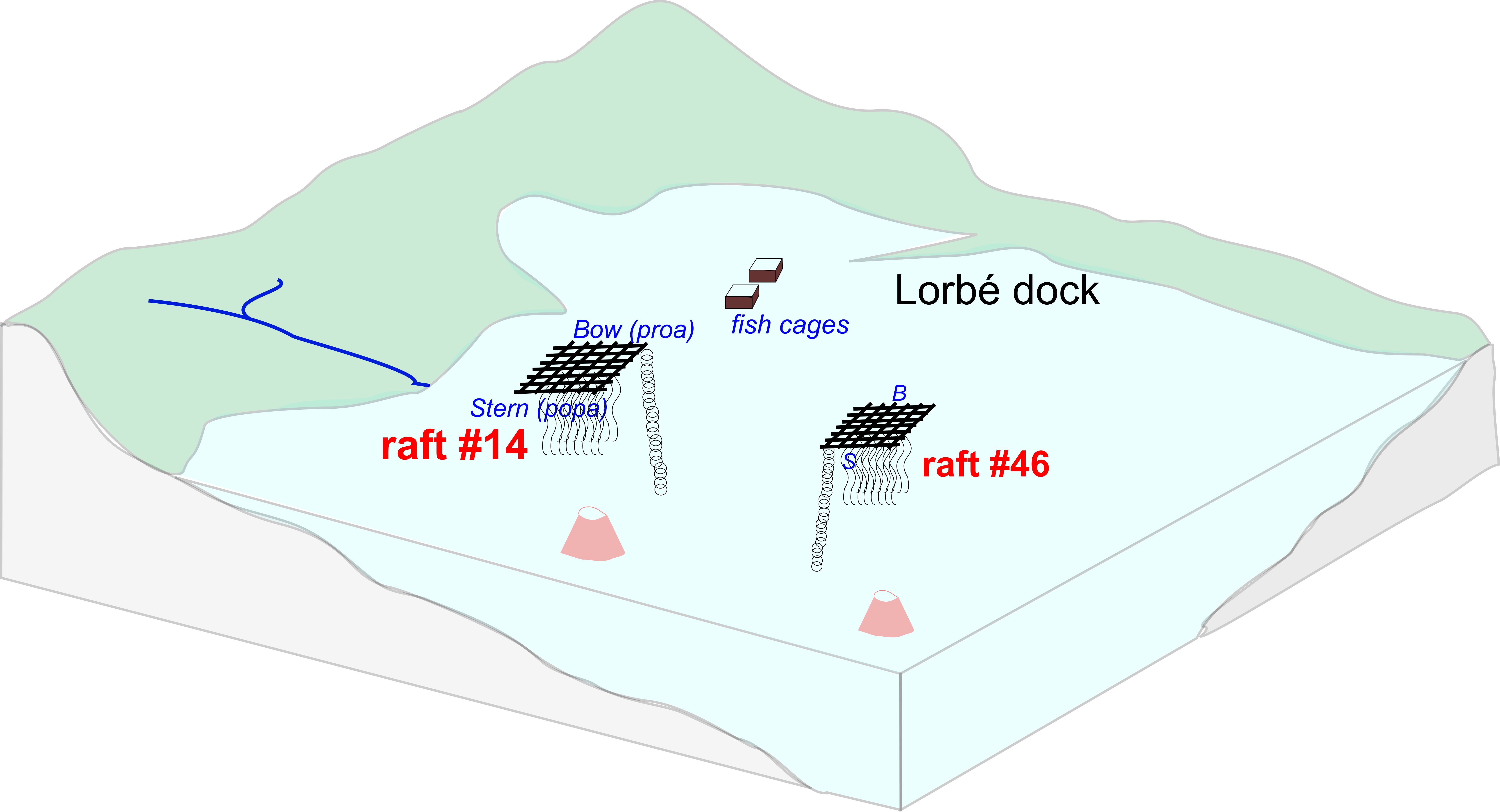
The Galician coastline is enormously productive of high-quality seafood due to the vast supply of nutrients, oxygen and plankton in this unique coastal upwelling system1. One of the characteristics of the Rías is the abundance of raft farms, “bateas”: floating wooden structures with 500 ropes hanging into the water column and on which mussels grow (suspended mussel farming). Galician mussel farms represent the highest mussel (Mytilus galloprovincialis) producing area in Europe (>250 Million kg per year)2. These farms are known to both boost biodiversity in the water column, providing refuge to fauna and anchoring for macroalgae, but also to cause alteration of benthic communities due to the massive production of “biodeposits” (mussel fall-out and faeces), sometimes resulting into anoxia at the seabed3,4. The Marine Research Institute (IIM) of the Spanish Research Council (CSIC) has been studying the ecology, chemistry and sustainability of these suspended bivalve systems for many years2,5. Now, an effort has been made to identify the potential of analytical pyrolysis techniques to provide information on biodeposit composition by means of analysis of particulate organic matter (POM) from sediment traps deployed underneath the farms.
Schematic representation of the analyzed system, showing the mussel cultures (rafts) and underlying sediment traps.
MATERIALS AND METHODS
The samples were obtained from a mussel culture polygon in the vicinities of Lorbé in the Ría de Ares-Betanzos (Sada, A Coruña province, Galicia, NW Spain), consisting of 107 rafts and an annual production of 10,000 tons of mussels. The particulate organic matter (POM) samples were obtained from multitrap collectors that were deployed beneath raft #46 (further from coast, 16 m water depth) and raft #14 (closer to coast and fish cages, 14 m depth) (Figure 1) on the 2nd and 3rd of May, 2011 (Spring bloom oceanographic scenario5). Samplers were deployed at the bow (or proa, i.e. water entry) and stern (popa, water exit) sides of the rafts. The traps contained four collectors which were open for no more than 12 h to mitigate the influence of microbial decomposition in the traps. Two of the 4 collectors within each trap were analyzed, T1 and T2 (these correspond to adjacent PVC cylinders within the multitrap). No preservatives were used (for details, see Zúñiga et al. 2014 5). Hence, a total of 8 samples were considered (2 rafts x 2 positions x 2 replicates) as a first approach to determine biodeposit sources and identify mechanisms that control variability in POM’s molecular composition. After retrieving the suspensions from the traps their content was filtered through Whatman GF/F filters (0.7 μm pore size). Due to the low inorganic matter content of these samples (<10 %), no sample treatment other than drying at 100 ºC, was performed (it would be a different scenario for the samples from Winter, which may have inorganic contents exceeding 75 %)5.
The Py-GC-MS analyses were performed by inserting solid POM (that was scraped directly from the filters) into quartz tubes with quartz wool on both ends. Pyrolysis was performed for 10 s at 650 °C, with a Pyroprobe 5000. The pyrolyzer was connected to a 6890 GC and 5975 MSD from Agilent Technologies. For chromatographic temperature profiles and MSD parameters, see Kaal et al. 6. Major peaks were quantified on the basis of characteristic m/z fragments. Data evaluation was based on i) pyrolysis compound percentages, analysis of variance (one-way ANOVA) and linear correlation analysis.
Table 1 presents the list of samples analyzed and parameters that are used for comparison with the Py-GC-MS data: fluxes of nitrogen (Ntotal), carbon (Ctotal) and organic C (Corg), which is modified data after Zúñiga et al. (2014)5.
Table 1. List of samples analyzed and selected parameters
Raft
Date
Sampler
Sample
Ntotal
Ctotal
Corg
m/d/yr
–
–
mg/m2d
mg/m2d
mg/m2d
#46 bow
5/2/2011
T1
17
405
2503
1603
5/2/2011
T2
18
304
1866
2092
#46 stern
5/2/2011
T1
19
813
5017
5101
5/2/2011
T2
20
619
4340
2833
#14 bow
5/3/2011
T1
27
186
1283
1045
5/3/2011
T2
28
210
1409
1321
#14 stern
5/3/2011
T1
29
447
3157
1958
5/3/2011
T2
30
488
3576
1930
RESULTS AND DISCUSSION
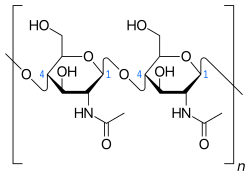
A large array of carbohydrate- and especially protein-derived pyrolysis products were detected (pyrroles, pyridines, diketodipyrrole, indoles) (Table 2). They include markers of N-acetylglucosamine polymers (acetamidosugars), which can be found in chitin from arthropod/crustacean/mollusk exoskeleta (and squid beaks), bacterial cell wall material, and the mucus of mollusks that cover faeces and pseudofaeces (Figure 2). The majority, if not all, of the other carbohydrate and protein products probably originate from mussel derivatives or marine biota such as phytoplankton.
Table 2. List of pyrolysis products and source allocation (CARB=carbohydrate, LIG=lignin, MAH=monocyclic aromatic hydrocarbon, MCC=metylene chain compound, NCOMP=N-compound, PAH=polycyclic aromatic hydrocarbon, PHEN=phenol)
#
RT (min)
compound
m/z
group
1
1.075
MeBr
94+96
other
2
1.371
benzene
78
MAH
3
1.569
pyridine
79+52
NCOMP
4
1.601
pyrrole
67
NCOMP
5
1.704
toluene
91+92
MAH
6
1.78
acetamide
59
NCOMP
7
1.897
3/2-furaldehyde
95+96
CARB
8
2.037
C1-pyrrole
80+81
NCOMP
9
2.146
C1-pyridine
93+66
NCOMP
10
2.396
styrene
104+78
MAH
11
2.786
5-methyl-2-furaldehyde
110+109
CARB
12
2.942
benzonitrile
103+76
NCOMP
13
3.082
phenol
94+66
PHEN
14
3.701
C1-phenol
107+108
PHEN
15
3.888
C1-phenol
107+108
PHEN
16
4.075
carbohydrate compound
126
CARB
17
4.569
C2-phenol
107+122
PHEN
18
4.845
naphthalene
128
PAH
19
5.115
propylcyanobenzene
91+131
NCOMP
20
5.193
4-vinylphenol
91+120
LIG
21
5.266
C3-phenol
121+136
PHEN
22
5.739
indole
117+90
NCOMP
23
5.843
3-acetamido-5-methylfuran
97+(69+)139
NCOMP
24
5.858
C1-naphthalene
142+141(+115)
PAH
25
5.864
p-tert-butylphenol
135+107(+150)
cont
26
5.931
C3:1 phenol (isopropenyl?)
134+119
cont
27
5.994
C1-naphthalene
142+141(+115)
PAH
28
6.139
3-acetamido-2/4-pyrone
111+(82+)153
NCOMP
29
6.55
C1-indole
130+131
NCOMP
30
6.566
biphenyl
154
PAH
31
8.214
fluorene
166+165
PAH
32
8.458
5-bromoindole
197+195(+116+89)
NCOMP
33
8.521
levoglucosan
60+73
CARB
34
8.988
diketodipyrrole
186+93
NCOMP
35
9.623
phenanthrene/anthracene
178
PAH
36
9.68
C14-fatty acid
60+73
MCC
37
10.117
alkane/alkene pair
55+57
MCC
38
10.158
phenylnaphthalene
204+203
PAH
39
10.283
phytadiene 1
68+95(278)
MCC
40
10.496
diketopiperazine
70+154+194
NCOMP
41
10.507
C16-alkylnitrile
97
NCOMP
42
10.579
phytadiene 2
81+82(278)
MCC
43
11.006
C16-fatty acid
60+73
MCC
44
12.191
C16-alkylamide
59+72
NCOMP
45
12.243
p,p‘-isopropylidenebisphenol
213+228
cont
46
12.243
C18-fatty acid
60+73
MCC
47
14.229
unknown compound
(91+)129+207
other
48
16.013
triterpenoid compound
368+353
other
49
16.663
triterpenoid compound
69+269(+298)
other
The most abundant group of products are the N-containing compounds, which account for 27.4 ± 3.1 % of TQPA (percentage of total quantified peak area), followed by monocyclic aromatic hydrocarbons (MAHs; 23.1 ± 3.7 %) and phenols (22.8 ± 5.1 %). This set of compounds is a clear indication that the POM originates from predominantly N-rich sources. The abundance of not only many protein markers (such as indoles from tryptophan or diketopiperazines from dimerization reactions of peptide chains during pyrolysis) but also products N-acetylglucosamine biopolymers suggests that mussel detritus, whether it be chitin from shells or mucus in faeces droppings, is a significant, if not dominant source of POM. Note that theoretically psuedofaeces (droppings of mucus-entangled inorganic matter filtered by the animal but which did not pass its digestive system) may also provide mucus to the biodeposits but due to the low seston concentration in the clear waters of NW Spain in general, production of pseudofaeces is unlikely (seston below threshold of 4 mg/L)5.
Fig. 2. Basic structure of N-acetylglucosamine polymers.
The proteins may originate from a combination of the mussel’s faeces, and alternative (marine) sources such as macroalgae and plankton. According to the high chlorphyll a content of these samples5, detritus from primary production is expected to be significant. The phenols may largely originate from aromatic amino acids in proteins such as tyrosine moieties, and the same can be argued for toluene among the MAH which is associated with phenylalanine7. Another component of the POM is of pyrogenic origin: products of e.g. charcoal from wildfires or soot from incomplete fuel combustion. This pyrogenic POM is reflected by PAHs (1.6 ± 0.4 % naphthalene, alkylnaphthalenes, fluorene, biphenyl, phenylnaphthalene, etc.), but also benzene (MAH) and benzonitrile5 (N-containing pyrogenic POM). This reaffirms previous indications5 of the presence of a minor fraction of allochthonous POM from runoff, with a high C/N ratio, and which is concentrated in the littoral areas in the studied ría 8. One sample produced a significant amount of a 5-methylbromoindole (sample 20, Table 1), whereas the pyrolyzates of the other samples contained trace amounts of this molecule. This may be a natural organobromine compound from marine biota9, or a secondary pyrolysis reaction between inorganic bromine and protein-derived indole, and has not been detected previously identified in POM pyrolyzates. Methyl-bromide (MeBr) is also abundant.
The long-chain aliphatic products (8.9 ± 1.9 %), or methylene chain compounds (MCC), also have various potential sources. Two phytadienes were detected among the MCC, which act as probably products of chlorophyll, from phytoplankton and/or macroalgae. The remaining products, notably fatty acids, may originate from any source but especially algae and mussel faeces were expected to be prolific of these compounds. Carbohydrate products account for 8.7 ± 2.3 %. They may be of any biological source mentioned above, but the high abundance of N-acetylglucosamine polysaccharide products among the N-compounds suggests that the carbohydrates may originate predominantly from chitin and/or mucus. Contamination indicators (4.9 ± 4.7 %) are tert-phenols and a bisphenol product. They probably both originate from bisphenol-type anthropogenic contamination. The C3:1-phenol compound shows similar differences in proportion among samples, and is therefore likely to originate from such contamination as well. Finally, lignin from terrestrial plants could not be unequivocally identified but the presence of a compound with m/z 120 and 91 at the expected retention time of 4-vinylphenol might reflect traces of land plant debris (if so, probably sedges or grasses including seagrasses).
For the ANOVA, we used all continuous variables as input attributes and the discrete parameters raft number (raft #46 vs. raft #14), trap position (bow vs. stern), and sampler (T1 vs. T2) as target variables.
As expected, none of the variables is affected by sampler number (T1/T2) located at the same spot (different cylinders in the same trap). For trap position, some significant (P<0.05) differences were observed. Firstly, the fluxes of Ctotal and Ntotal higher in the trap deployed at the stern position (popa). This suggests that the waters that flow through the rafts and the mussel cords gain in POM content along this path through the raft and illustrate biodeposition intensity differences. In other words, the effects of the rafts on the deposition of faeces can be observed not only from mass fluxes but also from relative contributions to POM. Among the Py-GC-MS products, stern samples are enriched in 3-acetamidopyrone (mussel chitin/mucus) and levoglucosan (chitin and perhaps other polysaccharides). Bow samples are enriched in MeBr. All the other >60 variables did not show significant differences.
For raft location, #14 was enriched in benzene, benzonitrile and fluorene (black-carbon derived) whereas #46 is enriched in some aliphatic products (an alkane/alkene pair and C16-alkylamide). This suggests that the site that is closer to the coastline is enriched in terrestrial black carbon materials, possibly indicative of fluxes of charcoal particles from rivers and streams that discharge in the area.
The linear correlations between the C and N fluxes showed some interesting relationships. Strong positive correlations between Ctotal and 3-acetamidopyrone (r=0.92!, P<0.005), C1-indole, diketopiperazines, levoglucosan and C16-fatty acid probably show the effect of POM sedimentation rate and proportions of mucus (delivering the N-acetylglucosamine polymer) and phytoplanikton (delivering protein). Negative correlations between Ctotal and MeBr, styrene and bisphenol products probably represents a dilution effect (less biodeposits, more background signal). The same patterns are observed for Corg and Ntotal. The C/N ratio is not correlated to any pyrolysis product.
Conclusions
The molecular properties of the POM could be largely traced back to nitrogen-containing biopolymers, in particular protein and mucus/chitin. Mucus and chitin cannot be distinguished on the basis of the available data yet, as we have not analyzed references materials for mussel mucus. The chitin/mucus originates predominantly from the mussel particles that trickle down from the rafts into the traps, possibly representing fall-off (although unlikely) and, most importantly, faeces. The protein originates from mussel debris (mucus/chitin) and plankton, and the presence of phytadienes provide strong evidence for algae-derived aliphatic POM (chlorophyll). The relative proportions of mucus/chitin and phytoplanktonic protein and aliphatic POM cannot be addressed: pyrolytic reactions are complex and chitin is entangled in protein which is difficult to distinguish from non-entangled protein in phytoplankton. However, from the ANOVA it was concluded that mussel debris was more abundant in the exit (stern) than entry (bow) of the mussel farms, which may reflect the background signal of POM from sources that are not directly associated to the bivalve cultures such as plankton. Other materials that were identified were traces of bisphenol contamination of unknown origin and fire residues (charcoal, black carbon), the latter of which had a stronger signal in the pyrolysis chromatograms from raft that was closest to the coastline (#14). In this context, it is worthy to mention that the region is subjected to a very intense fire regime. There is a clear potential of molecular characterization of particulate debris underneath mussel rafts to understand organic matter cycling and seafloor ecology.
ACKNOWLEDGEMENTS
The efforts of Uxío Labarta, María-José Fernández-Reiriz, Diana Zúñiga and Carmen González Castro, among other members of the CSIC-PROINSA Mussel Farm Team, allowed for the materialization of this collaboration.
References
Fraga, F., 1981. Upwelling off the Galician Coast, Northwest Spain. Book Series: Coastal and Estuarine Sciences, Volume 1: 176-182. https://doi.org/10.1029/CO001p0176.
Labarta, U., Fernández-Reiriz, M.J., Pérez-Camacho, A., Pérez-Corbacho, E., 2004. El mejillón, un paradigma bioeconómico. In: Fundación Caixa Galicia (Ed.), Bateeiros, mar, mejillón. Una perspectiva bioeconómica: Centro de Investigación Económica y
Financiera. Editorial Galaxia, Vigo, pp. 19–479
Otero, X.L., Calvo de Anta, R.M., Macías, F., 2008. Iron geochemistry under mussel rafts in the Galician ria system (Galicia-NW Spain). Estuarine and Coastal Shelf Science 81, 83–93.
Chamberlain, J., Fernandes, T.F., Read, P., Nickell. T.D., Davies, I.M., 2001. Impacts of biodeposits from suspended mussel (Mytilus edulis L.) culture on the surrounding surficial sediments. ICES Journal of Marine Science, 58: 411–416, doi:10.1006/jmsc.2000.1037.
Zúniga, D., Castro, C.G., Aguiar, E., Labarta, U., Figueiras, F.G., Fernández-Reiriz, M.J., 2014. Biodeposit contribution to natural sedimentation in a suspended Mytilus galloprovincialis Lmk mussel farm in a Galician Ría (NW Iberian Peninsula). Aquaculture 432, 311–320.
Kaal, J., Martínez Cortizas, A., Nierop, K.G.J., Buurman, P., 2008. A detailed pyrolysis-GC/MS analysis of a black carbon-rich acidic colluvial soil (Atlantic ranker) from NW Spain. Applied Geochemistry 23, 2395-2405.
Tsuge, S., Matsubara, H., 1985. High-resolution pyrolysis-gac chromatography if proteins and related materials. Journal of Analytical and Applied Pyrolysis 8: 49-64.
Sánchez-Mata, A., Glémarec, M., Mora, J., 1999. Physicochemical structure of the benthic environment of a Galician ría (Ría de Ares–Betanzos, north-west Spain). Journal of the Marine Biological Association of the United Kingdom 79, 1–21.
Andreotti, A., Bonaduce, I., Colombini, M.P., Ribechini, E., 2004. Characterisation of natural indigo and shellfish purple by mass spectrometric techniques. Rapid Communications in Mass Spectrometry 18: 1213–1220.
Itla-okla (Tillandsia usneoides) fibre temper in Pre-Columbian ceramics
By Joeri Kaal (Pyrolyscience) and Zackary Gilmore (Department of Anthropology, Rollins College, USA)
Zack Gilmore
Itla-okla, which means “tree hair”, was the Indian name of the epiphyte Tillandsia usneoides, also known as “Spanish beard” or “Spanish moss”. Pre-Columbian societies of the Late Archaic period (Orange and Stallings traditions; 3000-1800 B.C.) used these epiphytes as an organic temper in the production of pottery. In collaboration with Zackary Gilmore (Rollins College, Florida, USA), Pyrolyscience studied archaeological sherds of this fiber-tempered (FT) ware in order to identify (other) organic materials used for their fabrication and assess the production conditions of this extraordinary pottery (Gilmore, 2015).
Materials and Methods
The materials analyzed are (1) ceramics, ball-milled to powder and treated with HF to eliminate oxides, (2) isolated charred fiber materials, scraped from the archaeological sherds and (3) Spanish moss, both uncharred and charred in a temperature series between 300 and 600 ºC. The analysis of charred “moss” (it is neither a moss nor a lichen, but an angiosperm from the Bromeliaceae family) was thought to enable the estimation of the firing temperature (in terms of muffle furnace equivalent temperature, TMFE) of the isolated fibres and whole ceramic fragments. HF treatment was performed to eliminate reactive minerals. Pyrolysis-GC-MS was performed at 750 ºC to stimulate fragmentation of thermoresistant components such as charred organic matter (Black Carbon) (Kaal et al., 2009). Extant moss was double-wrapped in aluminum foil to create oxygen-limited conditions and charred between 300 and 600 ºC (e.g. Turney et al., 2006).
Results and Discussion
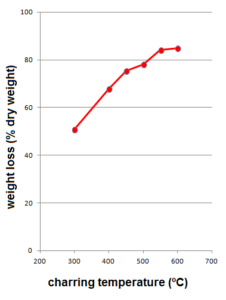
First we’ll discuss the molecular chemistry of the plant and it’s experimentally created charred equivalents. The loss of weight of the sample of fresh Itla-okla as induced by charring in the muffle furnace shows a typical increase from low to high temperatures: 50 % at TMFE=300 ºC towards 85 % at TMFE=600 ºC (Figure 1). This is known to reflect dehydration and condensation reaction as the temperature rises.
Figure 1. Weight loss plotted against charring temperature (muffle furnace experimental heating)
With Py-GC-MS, the uncharred sample (feedstock) is prolific of many polysaccharide products (acetic acid, furans, cyclopentenones, pyranones), lignin products (4-vinylphenol, guaiacols, syringol) and some aliphatic products such as phytadienes (from chlorophyll) and fatty acids. With THM-GC-MS, the main peaks are fatty acid methyl esters (C16, C18, C24-C28), mid-chain methoxylated C16 fatty acid methyl esters (from cutin in cuticula) and the lignin products P18 (p-coumaric acid methyl ester), G18 (ferulic acid methyl ester). These results represent the first molecular screening with Py-GC-MS and THM-GC-MS of Tillandsia usneoides feedstock.
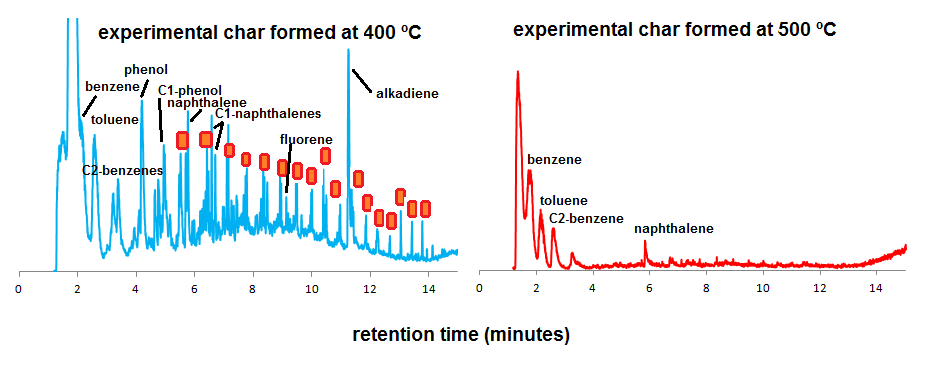
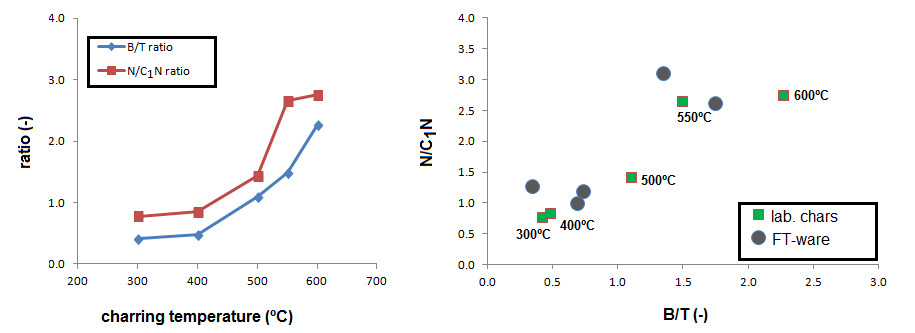
With increasing TMFE, the peak intensities (Py-GC-MS only) of these polysaccharide and lignin products decrease and those of monocyclic and polycyclic aromatic hydrocarbons (MAHs/PAHs) increase, which is a feature that has been observed in many kinds of feedstock materials of which experimental “thermosequences” were analyzed by Py-GC-MS. There is an intermediate range of thermal modification as recognized from Py-GC-MS fingerprints between 300 and 400 ºC with abundance of aliphatic products (alkanes, alkenes) and phenols (degraded lignin). The abundance of these phenols and aliphatic products (n-alkanes and n-alkenes from aliphatic biopolymers such as cutin) is high in the samples produced at 300 and 400 ºCMFE but not at 500 ºC and higher (here, the dominance of MAHs and PAHs is very strong) (Figure 2). These results are in agreement with previous studies of plant-derived chars.
The relative abundances of benzene and toluene (B/T ratio) and naphthalene and methylnaphthalens (N/C1N ratio) has been used as an indicator of charring intensity. The graph below shows how these parameters increase with TMFE in the experimental graph, especially between 400 and 550 ºC. These trends can be used to compare with the fibres and whole sherds from the FT ceramics. The figure to the right shows the B/T – N/C1N plot of the laboratory chars and the isolated fibers, and shows that three of the fibers plot in the low-thermal impact range (<500 ºCMFE) whereas the other two plot in the high temperature range (450–600 ºCMFE). For the whole sherds, we could not establish B/T due to the existence of double peaks (implying a combination of volatiles and polymeric sources) and the methylnaphthalenes could not be reliably quantified due to low peak intensities.
Figure 3. Benzene/toluene and napththalene/methylnaphthalenes ratio. Left figure: as a function of charring temperature for laboratory chars. Right figure: ratios plotted for both laboratory chars of Spanish moss and pottery fragments.
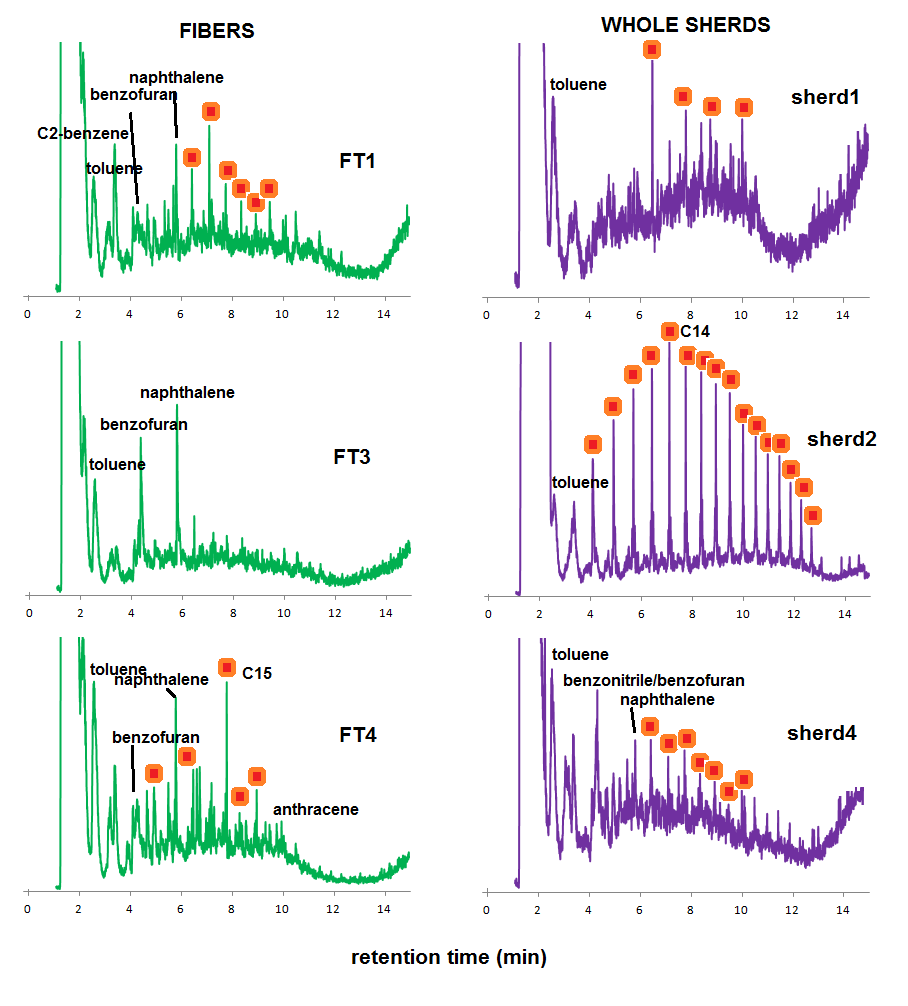
This information can now be used to interpret the Py-GC-MS fingerprints of the fibers extracted from sherds and whole sherd samples (Figure 4). From the chromatograms of the isolated fibers (charred elements in sherds), there are clear differences in thermal impact, which were also observed by tracking the B/T ratio (Figure 3, right graph). Some samples are dominated by MAHs and PAHs, and benzofuran, such as sample FT3 in Figure 4. Others have higher proportions of aliphatic compounds, such as FT1 and FT4, indicative of lower thermal impact.
Figure 4. Py-GC-MS chromatograms of Spanish moss fibers isolated from pottery (green, left) and whole sherd materials (purple, right). Red/orange symbols represent alkane/alkene pairs.
The chromatograms of whole sherds are more surprising. Sample sherd2, for example, shows a pyrolysis fingerprint that is strongly dominated by alkanes and alkenes, and with a chain length pattern that is clearly distinct as what was observed for the experimental chars or the isolated fibers from the archaeological sherds. This may be indicative of other organic ingredients (bitumen, dung or plant materials, for example) that were added to the ware before firing and were absorbed to it during its use. Sherd4 has a fingerprint that can be largely ascribed to Itla-okla remains.
Conclusions
Clearly, there are differences in firing intensity as observed from the balance between aliphatic products and phenols, vs. MAHs and PAHs. This became evident by comparing the Py-GC-MS fingerprints of the experimental thermosequences and the archaeological ware (isolated fibers). The range of temperatures are <500 and 500-600 ºCMFE. Note that these differences do not necessarily imply that there are large differences in the maximum temperature during firing: other factors such as the thickness of the material, and the duration of the process can explain the results as well. For example, relatively short firing or the use of thick ware may cause incomplete burn-off in the center of the ware where the organic matter may be least affected by the thermal alteration: such features can be recognized as sandwich morphologies of sherds (see figure below: dark core, lighter interior and exterior surfaces). The results of the whole sherd analysis may be indicative of other organic constituents and future studies will focus on the characterization of those materials.
References
Gilmore, Z.I., 2015. Direct radiocarbon dating of Spanish moss (Tillandsia usneoides) from early fiber-tempered pottery in the southeastern U.S. Journal of Archaeological Science 58, 1–8.
Kaal, J., Nierop, K.G.J., Martínez Cortizas, A., 2009. Characterisation of aged charcoal using a coil probe pyrolysis-GC/MS method optimised for Black Carbon. Journal of Analytical and Applied Pyrolysis 85, 408-416.
Kaal, J., Lantes Suárez, O., Martínez Cortizas, A, Prieto, B., Prieto Martínez, M.P., 2014. How useful is pyrolysis-GC-MS for the assessment of molecular properties of organic matter in archaeological pottery matrix? An exploratory case study from NW Spain. Archaeometry 56, 187-207.
Turney, C.S.M., Wheeler, D., Chivas, A.R., 2006. Carbon isotope fractionation in wood during carbonization. Geochimica et Cosmochimica Acta 70, 960–964.
Posidonia oceanica meadow in the Mediterranean Sea. Photo: Eduard Serrano
A recent paper in Organic Geochemistry entitled “Radically different lignin composition in Posidonia species may link to differences in organic carbon sequestration capacity” discusses the remarkable difference in lignin chemistry between two kinds of “Neptune grass”, i.e. Posidonia oceanica and Posidonia australis.
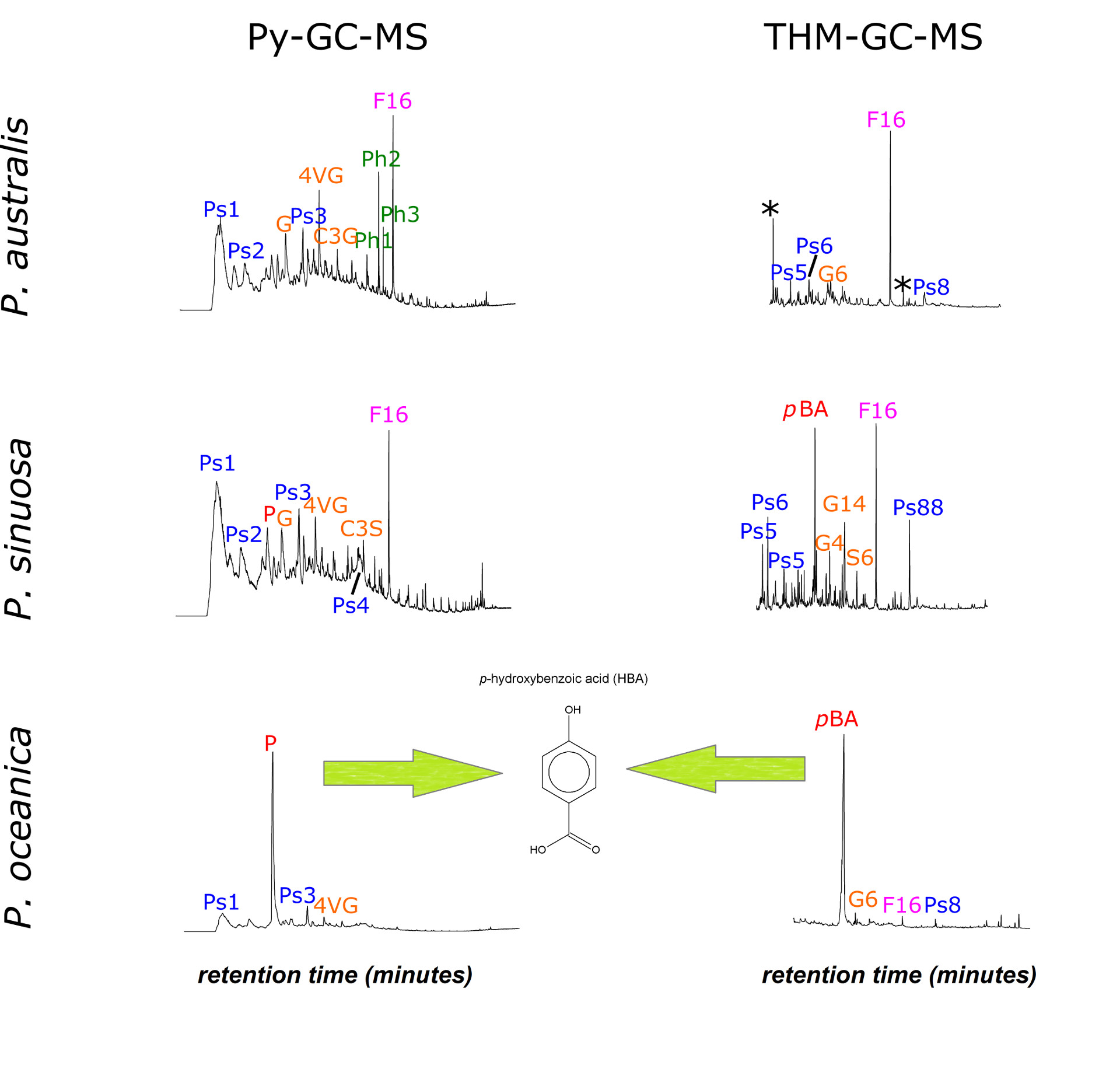
Initial efforts using analytical pyrolysis techniques (Py-GC-MS and THM-GC-MS) showed that the endemic Mediterranean member of the Posidonia genus, P. oceanica, has abnormally high amounts of p-HBA (para-hydroxybenzoic acid) whereas the down-under variety does not. State-of-the-art lignin characterization in Seville (DFRC, 2D-NMR) showed that the p-HBA is part of the lignin backbone, and not glycosylated as initially expected, and P. oceanica is now the producer of the most extensively p-HBA-acylated lignin known in the Plant Kingdom.
Chiefly the Mediterranean Posidonia (=P. oceanica) creates massive submarine peat deposits called mats. And only P. oceanica has p-HBA-burdened lignin. The paper in Organic Geochemistry discusses the likeliness of the peculiar lignin structure as the main cause of the differences in their potential to accumulate debris in mats: microbes would not be able to decompose it.
Posidonia oceanica mat deposits in Portlligat (Western Mediterranean). Photo: Kike Ballesteros
Here we will have a look at a bit of unpublished data on a third Posidonia species. More specifically, we discuss the composition of the sheath materials of Posidonia sinuosa, which is endemic to western and southern Australia. Contrary to P. oceanica and P. australis, for this species the molecular characterization was not performed for a range of anatomical sections of the seagrass (leaf blade, leaf sheath, rhizome, root) but for the leaf sheath only. This is the part where lignin is concentrated and therefore its analysis is thought to be representative of the lignin features of this plant species.
From the Py-GC-MS and THM-GC-MS data, the P. sinuosa is much more similar to its southern relative that to the Mediterranean high-carbon accumulator (see figure below), but it probably contains slightly more p-HBA than Posidonia australis. Posidonia sinuosa is not capable of producing big mat deposits neither, and it is tempting to conclude that there is a link between the palatability of different species of Posidonia seagrasses (by microbes, that is) and the abundance of p-HBA in the lignin backbone, even though definitive evidence has yet to be presented.
In a similar fashion as land plants, Posidonia species are major sequesters of atmospheric carbon. Considering that they also manage to store it in millenary years old sea-peat deposits, storing many gigatons of carbon that would otherwise raise CO2 pressures, it is not surprising that these seagrass habitats, which are, despite of the numerous governmental entities that have developed programs for their protection -including cultivation programs-, in dire need of conservation.
Even though the Australian and Mediterranean genera diverged tens of Millions ago (possible associated with the closure of the Tethys Sea), it is striking that members of the same family can have such a remarkable difference in the structure of their skeletons: lignin is not a minor chemical but it accounts for up to 30 % of the seagrasses dry weight and is the largest biomolecular constituent after holocelluloses.
Pyrolysis-GC-MS and THM-GC-MS total ion current chromatograms of P. australis, P. sinuosa and P. oceanica, showing that only P. oceanica is prolific of p-HBA derivatives upon analytical pyrolysis (but P. sinuosa has more than P. australis). Symbols: F16= C16-fatty acid, G= guaiacols, P= phenol, Ps= polysaccharide, Ph1= isoprenoid chlorophyll product, pBA= p-hydroxybenzoic acid methyl ester, * = contamination (TMAH products and column bleed).
Bibliography:
Aires, T., Marbà, N., Cunha, R.L., Kendrick, G.A., Walker, D.I., Serrão, E.A., Duarte, C.M., Arnaud-Haond, S., 2011. Evolutionary history of the seagrass genus Posidonia. Marine Ecology Progress Series 421, 117–130. doi: 10.3354/meps08879
Fourqurean, J.W., Duarte, C.M., Kennedy, H., Marba, N., Holmer, M., Mateo, M.A., Apostolaki, E.T., Kendrick, G.A., Krause-Jensen, D., McGlathery, K.J., Serrano, O., 2012. Seagrass ecosystems as a globally significant carbon stock. Nature Geoscience 5 (7), 505–509.
Kaal, J., Serrano, O., Nierop, K.G.J., Schellekens, J., Martínez Cortizas, A., Mateo, M.-A., 2016. Molecular composition of plant parts and sediment organic matter in a Mediterranean seagrass (Posidonia oceanica) mat. Aquatic Botany 133, 50-61.
Kaal , J., Serrano, O., Del Río, J.C., Rencoret, J., 2018. Radically different lignin composition in Posidonia species may link to differences in organic carbon sequestration capacity. Organic Geochemistry 124, 247-256.